
Written By: Xiu Li
MB0902
092908H
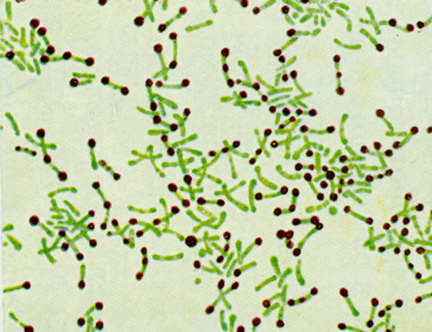
A pilus is a hairlike appendage found on the surface of many bacteria. The term pilus and fimbriae are used interchangeably, although some researchers reserve the term pilus for the sexual appendage required for bacterial conjugation.MB0902
092908H
Pili connect a bacterium to another of its species, or to another bacterium of a different species, and build a bridge between the cytoplasm of the cells. This enables the transfer of plasmids between the bacteria, an exchanged plasmid can code for new functions. The pilus is made out of the protein flagellin.
Up to ten of these structures can exist on the bacteria. Some bacterial viruses or bacteriophages attach to receptors (a protein molecule embedded in either the plasma cembrane or cytoplasm of a cell) on sex pili at the start of their cycle.
Sex pili
A pilus is typically 6 to 7nm in diameter.
Despite its name, the sex pilus is not used for sexual reproduction.
During bacterial conjugation, a sex pilus emerging from one bacterium ensnares the recipient bacterium, draws it in, and eventually triggers the formation of a mating bridge. This establishes direct contact, merging the cytoplasms of two bacteria via a controlled pore. This pore allows for the transfer of bacterial DNA from the bacteria with the pilus (donor) to the recipient bacteria.

Through this mechanism of genetic transformation, advantageous genetic traits can be disseminated amongst a population of bacteria. Not all bacteria have the ability to create sex pili, however sex pili can form between bacteria of different species.

The fertility factor is required to produce a sex pili.
(A bacterial DNA sequence that allows a bacterium to produce a sex pilus necessary for conjugation)
Fimbriae
A fimbria is a a proteinaceous appendage in many gram-negative and gram-positive bacteria that is thinner and shorter than a flagellum. This appendage ranges from 3-10 nanometers in diameter and can be up to several micrometers long. In Gram positive bacteria, the pilin subunits are covalently linked.
Fimbriae are used by bacteria to adhere to one another and to adhere to animal cells, and some inanimate objects. Attachment of bacteria to host surfaces is required for colonization during infection or to initiate formation of a biofilm. Mutant bacteria that lack fimbriae cannot adhere to their usual target surfaces and, thus, cannot cause diseases.
Fimbriae are either located at the poles of a cell, or are evenly spread over its entire surface. A bacterium can have as many as 1,000 fimbriae.
Fimbriae are only visible with the use of an electron microscope.






































{kind=link}
{kind=link}
{kind=link}